| This page in English! |
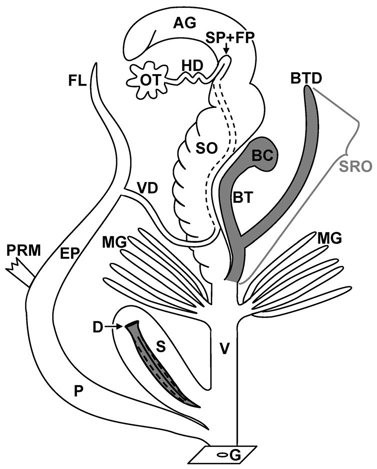 Schematische Abbildung des Genitalapparats einer gefleckten Weinbergschne- cke (Cornu aspersum). Quelle: Koene, Schulenburg: BMC Evolutionary Bio- logy 2005, 5:25. |
Das Organsystem des Genitalapparats im Körper einer Weinbergschnecke enthält nicht nur Geschlechtsorgane im eigentlichen Sinn, sondern auch unterschiedliche Hilfsorgane, die in verschiedenen Phasen der Paarung Aufgaben zu erfüllen haben. Die Geschlechtsöffnung befindet sich auf der rechten Körperseite kurz hinter dem Kopf. Von hier aus reichen die Organe des Genitalapparats bis tief in den Eingeweidesack.
Bei den seltenen links gewundenen Mutanten der Weinbergschnecke ("Schneckenkönige") öffnet der Genitalapparat sich stattdessen auf der linken Körperseite.
Auf den ersten Blick erscheint der Genitalapparat der Weinbergschnecke sehr kompliziert gebaut zu sein. Daher sollen hier die einzelnen Organe und Organgruppen auch in Zusammenhang mit ihrer Rolle während der einzelnen Phasen der Paarung betrachtet werden.
Zwittrige Bereiche:
AG: Eiweißdrüse; OT: Zwitterdrüse; HD: Zwittergang; SP + FP: Samentasche (Spermatheca)
und Befruchtungstasche; SO: Eisamenleiter; G: Atrium.
Weibliche Bereiche:
BC: Bursa copulatrix; BTD: Divertikel; MG: Fingerförmige Drüsen; V: Vagina;
S: Pfeilsack; D: Liebespfeil.
Männliche Bereiche:
VD: Vas deferens (Samenleiter); FL: Flagellum; EP: Epiphallus; P: Penis; PRM:
Penisrückziehmuskel.
Die Keimzellen der Weinbergschnecke, Samen- und Eizellen, entstehen in einer gemeinsamen, zwittrigen Keimdrüse oder Gonade, der Zwitterdrüse. Eizellen und Samenzellen entstehen prinzipiell auf dieselbe Weise, indem Wandzellen der Zwitterdrüse zu Mutterzellen ausdifferenzieren, aus denen sich nach mehreren Zellteilungen (Mitosen) und einer Reifeteilung (Meiose) über mehrere Zwischenstadien schließlich die endgültigen Keimzellen bilden. Allerdings entstehen im Gegensatz zu vielen Samenzellen, wie in der Natur allgemein üblich, nur einige wenige Eizellen.
Während die Eizellen erst kurz vor der Befruchtung die Zwitterdrüse verlassen, werden Samenzellen laufend in den Zwittergang abgegeben. Dort werden sie in taschenförmigen Aussparungen, den Samenvesikeln, in unbeweglichem Zustand gespeichert, bis sie zum Einsatz kommen. Dann wandern sie aus eigener Kraft aus dem Zwittergang in den Eisamenleiter (Spermovidukt) aus.
Im Eisamenleiter ist der Samenleiter (Spermidukt) als rinnenförmiger Kanal offen mit dem Eileiter verbunden, so dass Samenzellen aus dem Samenleiter in den Eileiter gelangen können. Während der Kopulation wird diese Verbindung jedoch geschlossen, so dass die Samenzellen nur durch den Samenleiter wandern können. In den Samenleiter mündet auch eine Drüse, die man als Spermiduktdrüse oder auch als Prostata bezeichnet. Sie gibt ein nährstoffreiches Sekret in den Samenleiter ab, das die Samenzellen ernährt und mit Energie versorgt. Da das Sekret der Prostata die Samenzellen außerdem inaktiviert, werden sie von hier aus nicht mehr aus eigener Kraft, sondern nur noch durch peristaltische Bewegungen der Kanalwände weiter transportiert. Weiter unten spaltet sich der Eisamenleiter in den Eileiter (Ovidukt) auf, der schließlich in die Vagina mündet, sowie in den Samenleiter (Vas deferens, wörtlich: ausführender Gang), der in den so genannten Epiphallus eintritt und durch den die Samenzellen in der Prostata-Flüssigkeit weitertransportiert werden.
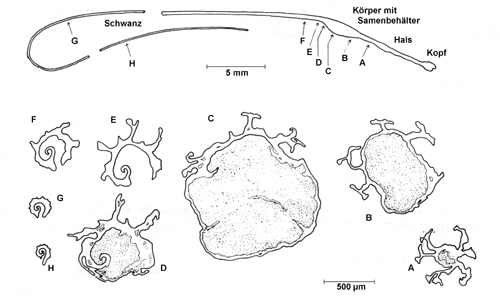 Die Spermatophore. An den Querschnitten D bis G ist gut zu erkennen, dass der Schwanz der Spermato- phore nicht geschlossen ist, so dass die Samenzellen nach der Übertragung auf diesem Weg entkommen können. Quelle: Lind (1973). |
Wie bereits erwähnt, findet die Übertragung der Samenzellen während der Kopulation der Weinbergschnecke mittels einer so genannten Spermatophore statt, einem spezialisierten Transportmittel. Während die Samenzellen heran transportiert werden, entsteht die Spermatophore im Bereich der Mündung des Vas deferens in den Epiphallus. Drüsenzellen im Epiphallus scheiden Baumaterial in den Hohlraum des Kanals aus, das an den Wänden erstarrt und daher ihre Oberflächenstruktur annimmt. Auf die Mündung des Vas deferens zu bleibt die entstehende Struktur jedoch offen.
Während sie nach vorne geschlossen ist, da hier Baumaterial von allen Seiten in den Kanal abgegeben wird, setzt sie sich nach hinten in das wegen seiner peitschenähnlichen Form so genannte Flagellum fort. Hier entsteht ein langer, fadenförmiger Schwanzfortsatz, der sich an den hohlen Körper der Spermatophore anschließt. Da das Flagellum eine längs verlaufende Bodenfalte aufweist, ist der Schwanz der Spermatophore nach unten offen.
Sobald die Masse der Prostataflüssigkeit mit den darin schwimmenden Samenzellen aus der Öffnung des Vas deferens austritt, bewegt sich die Spermatophore nach vorne. Dadurch wird schließlich die Öffnung hinter den Samenzellen geschlossen. Ganz vorne wird der geschlossene Teil der Spermatophore durch Kontraktionen der Peniswand zu einer Kopf-Nacken-Struktur verformt.
Im Bild oben sollen sich die Querschnitte G und H 4 bzw. 8 cm vom eigentlichen Körper der Spermatophore entfernt befinden. Die gesamte Spermatophore einer Helix pomatia einschließlich Schwanz ist also etwa 11 cm lang.
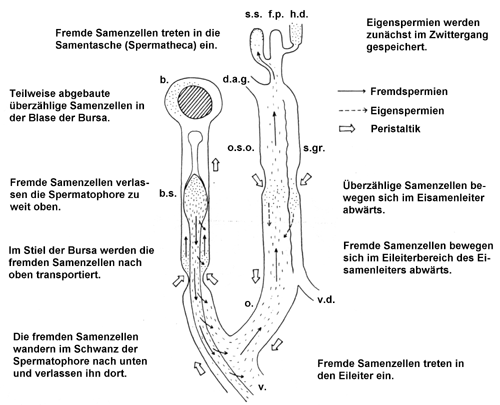 Schema: Der Weg der Samenzellen nach der Kopulation. Beschriftung: b.: Blase der Bursa copulatrix; b.s.: Stiel der Bursa; d.a.g: Kanal der Eiweißdrüse; s.s.: Samentasche; f.p.: Befruchtungstasche; h.d.: Zwittergang; o.s.o.: Ovidukt-Teil des Spermoviduktes; s.gr.: Samenrinne (Spermidukt); o: Eileiter (Ovidukt); v.d.: Vas deferens. Quelle: Lind (1973). |
Die Spermatophore mit den darin eingelagerten Samenzellen wird nun mit dem Penis in den Genitalapparat des Partners übertragen. Dort wird sie in einem Seitenkanal der Vagina platziert. Dieser Kanal bildet zusammen mit seinem blasenförmigen Endstück die Bursa copulatrix (Begattungstasche).
Die Bedeutung der Bursa copulatrix war lange Zeit unbekannt. Man ging davon aus, sie diene der Aufnahme und Speicherung der fremden Samenzellen. Daher findet man vielfach die Bezeichnung Receptaculum seminis (wörtlich "Samenempfänger").
Tatsächlich ist die Rolle der Bursa copulatrix eine völlig andere: In der Endblase der Bursa werden Samenzellen durch Enzyme zerlegt und abgebaut, nicht gespeichert.
Vor diesem Hintergrund dient die Spermatophore nicht nur als Übertragungsmedium, sondern auch dem Schutz der darin enthaltenen Samenzellen. Während nämlich der Kopf der Spermatophore unaufhaltsam durch peristaltische Bewegungen des Bursa-Stiels in Richtung Endblase bewegt, können die Samenzellen aus dem Samenbehälter in den Schwanz der Spermatophore entkommen. Da dieser, wie bereits beschrieben, nicht geschlossen ist, entkommen die Samenzellen hier nach außen.
Die peristaltischen Bewegungen des Bursa-Stiels würden dennoch dazu führen, dass die Samenzellen außerhalb der Spermatophore in die Endblase befördert werden und dort der Vernichtung anheim fallen. Der Schwanz der Spermatophore ist mit fast 10 cm Länge jedoch deutlich länger als der Stiel der Bursa, so dass einige Samenzellen bei ihrem Austritt fast an die Mündung des Eileiters gelangen.
Trotz des spezialisierten Baus der Spermatophore schaffen im Normalfall nur ein sehr geringer Anteil, etwa 1‰, der übertragenen Samenzellen den Eintritt in den Eileiter.
Welchen biologischen Sinn hat nun ein Organ, wie die Bursa copulatrix, das die mühsam nach mehreren Stunden Paarung übertragenen Samenzellen vernichtet? Dazu man muss sich vor Augen führen, dass im Rahmen der geschlechtlichen Fortpflanzung immer wenige eigene Eizellen, manchmal sogar nur eine, einer Vielzahl fremder Samenzellen gegenüberstehen. Angesichts dessen liegt es natürlich im Interesse des Empfängers der Samenzellen, nur wenige, aber dafür die leistungsfähigsten, Samenzellen des Partners zu erhalten, um damit schließlich die wenigen wertvollen Eizellen zu befruchten. Die Bursa copulatrix selektiert daher aus der Vielzahl der übertragenen Samenzellen die wenigen aktivsten aus, die ihr entkommen können. Neben Fremdspermien und Spermatophoren baut die Bursa copulatrix auch überzählige eigene Samenzellen und Prostataflüssigkeit ab. Aus diesem Grund ist die Verbindung zwischen Samenleiter und Eileiter im Eisamenleiter außerhalb einer Kopulation offen, wird bei der Kopulation jedoch geschlossen, so dass die eigenen Samenzellen nicht etwa in der eigenen Bursa copulatrix enden, sondern möglichst alle in der Spermatophore übertragen werden.
Gegenüber der Bursa copulatrix des Empfängers bleibt aber auch dem Spender der Samenzellen eine Möglichkeit, die Verhältnisse zu seinen Gunsten zu beeinflussen: Das mit dem Liebespfeil übertragene Sekret der fingerförmigen Drüsen enthält Hormone, die den Stiel der Bursa und den Eileiter beeinflussen. So wird der Bursa-Stiel erweitert, so dass die Samenzellen leichter entkommen können. Der Eileiter wird in peristaltische Bewegungen versetzt, von denen die Eigenbewegung der Samenzellen unterstützt wird.
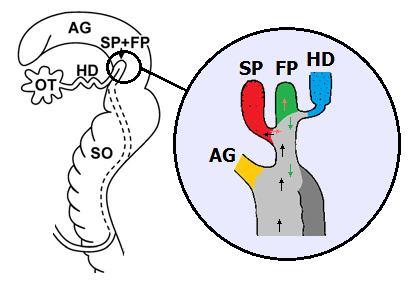 Samentasche (SP) und Befruchtungstasche (FP) bei Cornu aspersum. Links: Lage im Genitalapparat. Vergrößerte Darstellung: Gelb: Gang der Eiweißdrüse. Rot: Samentasche. Grün: Befruchtungstasche. Blau: Zwittergang. Weg der Samenzellen: Schwarz: Auf dem Weg zur Samentasche. Hellrot: Von der Sa- mentasche zur Befruchtungstasche. Weg der befruchteten Eizellen: Grün. Hellgrau: Eisamenleiter; Dunkelgrau: Samenleiter (nur teilweise getrennt). Weitere Abkürzungen: s.o. Quellen: Koene, Schulenburg: BMC Evolutionary Biology 2005, 5:25 (links) und Lind (1973) (rechts). Überarbeitet von R. Nordsieck. |
Durch den Eileiter gelangen die wenigen überlebenden Samenzellen bis zum Ende des Eisamenleiters. Hier befindet sich, eingelagert in die große Eiweißdrüse, die Befruchtungstasche der Schnecke. Ein abgegrenzter Blindsack der Befruchtungstasche dient als Samentasche (Spermatheca). Hier werden die Samenzellen aller Paarungspartner einer Schnecke, die den Weg bis hier geschafft haben, gespeichert. Zur Befruchtung müssen sie schließlich nur noch den kurzen Weg bis in die Befruchtungstasche der Schnecke zurücklegen.
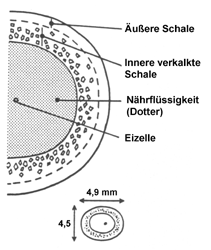 Querschnitt durch ein Ei von Cornu asper- sum. Quelle: Tompa (1984). |
Erst in unmittelbarem Zusammenhang mit der Eiablage werden die Eizellen aus der Zwitterdrüse in die Befruchtungstasche der Weinbergschnecke transportiert, so dass es dort zur Befruchtung durch die Samenzellen aus der Samentasche kommen kann. Von der Eiweißdrüse werden die befruchteten Eizellen dann mit einem nährstoffreichen Hülle versehen, die entgegen des Namens der Drüse nicht nur aus Eiweiß, sondern auch aus Glykolipiden und anderen Nährstoffen besteht.
Die noch schalenlosen Eier werden von hier aus in den Eileiter transportiert, wo sie sich bis zu ihrer Ablage wie Perlen an einer Schnur aufreihen. Man bezeichnet diesen Teil des Eileiters einer Weinbergschnecke auch als Uterus, denn Drüsenzellen in der Wand des Eileiters umhüllen die Eier mit mehreren Eihüllen, die teilweise stark verkalkt werden und so eine Eierschale bilden. Wie man bei einer Durchleuchtung des Uterus erkennen kann, findet die Verkalkung der Eierschalen graduell statt, so dass die am oberen Ende des Uterus liegenden Eier deutlich durchsichtiger sind, als die nahe dem Ausgang.
Zusätzlich zu ihrer Kalkschale werden die Eier der Weinbergschnecke hier auch mit einer Schleimschicht umhüllt, die sie in der Legehöhle vor Pilzbefall und anderen Infektionen schützt.
Zum Zeitpunkt ihrer Ablage haben die 40 – 60 Eier einer Weinbergschnecke einen Durchmesser von 7 – 8 mm erreicht.